MAXAM-GILBERTOVA METODA
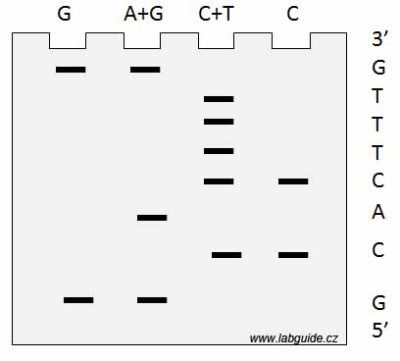
Tato metoda využívá chemického štěpení jednotlivých typů bází. Metoda pracuje s jednovláknovou DNA, která je na svém jednom konci (3’ nebo 5’) radioaktivně značena. Reakce je prováděna ve čtyřech zkumavkách, přičemž v každé zkumavce je prováděno štěpení jen určitých typů bází. To znamená, že DNA se štěpí jen v místě určitých bází. Tím vzniká směs různě dlouhých fragmentů, které končí v místě určité báze a jejich elektroforézou v hustém polyakrylamidovém gelu, kdy jsou všechny čtyři reakce naneseny vedle sebe, určíme rozdíly v délce fragmentů, a tedy i jak daleko od začátku fragmentu tato báze byla. Odečtením pozice jednotlivých bází ve všech čtyřech reakcích stanovíme sekvenci daného úseku.
SANGEROVA METODA
Sangerova metoda využívá proces replikace DNA a tedy princip využívaný také při PCR. Původní a klasické provedení metody je prováděno tak, že:
• k jednořetězcové DNA přisedá 15-25 bp dlouhý primer (značený radioaktivně), který je komplementární k začátku sekvenovaného místa,
• od navázaného primeru probíhá syntéza DNA za přítomnosti dNTP (dATP, dCTP, dGTP a dTTP) a jednoho z dideoxynukleotidů (ddATP, ddCTP, ddGTP nebo ddTTP). Jednotlivé dideoxynukleotidy jsou v porovnání s jednotlivými nukleotidy v reakci zastoupeny jen v relativně malém množství,
• reakce je prováděná ve čtyřech zkumavkách, kdy každá ze zkumavek obsahuje „svůj“ dideoxynukleotid (ddNTP),
• dideoxynukleotidy se náhodně začlení do syntetizovaného řetězce místo příslušného dNTP (např. místo dATP dosedne ddATP). A protože ddNTP nemají OH skupinu, po jejich dodání do syntetizovaného řetězce se syntéza zastaví. Vzhledem k tomu, že v každé reakci je obrovské množství molekul DNA a že začleňování ddNTP se děje náhodně, a to díky jejich nízké koncentraci jen s nízkou pravděpodobností (část molekul je v příslušném místě obsazena dNTP a část ddNTP), vzniká v každé reakci směs různě dlouhých fragmentů. Délka každého z fragmentů udává pozici příslušného ddNTP,
• délka fragmentů je analyzovaná pomocí elektroforézy v polyakrylamidovém gelu, a to tak, že se na gel nanesou vedle sebe produkty ze všech čtyř zkumavek a po separaci fragmentů se díky radioaktivnímu značení primerů odečte délka jednotlivých fragmentů. Pozice jednotlivých fragmentů odpovídají pozicím jednotlivých nukleotidů v sekvenovaném vzorku.
V modifikované podobě Sangerova metoda využívá fluorescenčně značených dideoxynukleotidů (každý ddNTP nese svou barevnou značku), což umožňuje provedení reakce v jedné zkumavce. Produkty jsou pak analyzovány kapilární elektroforézou a je získán výsledný sekvenogram.
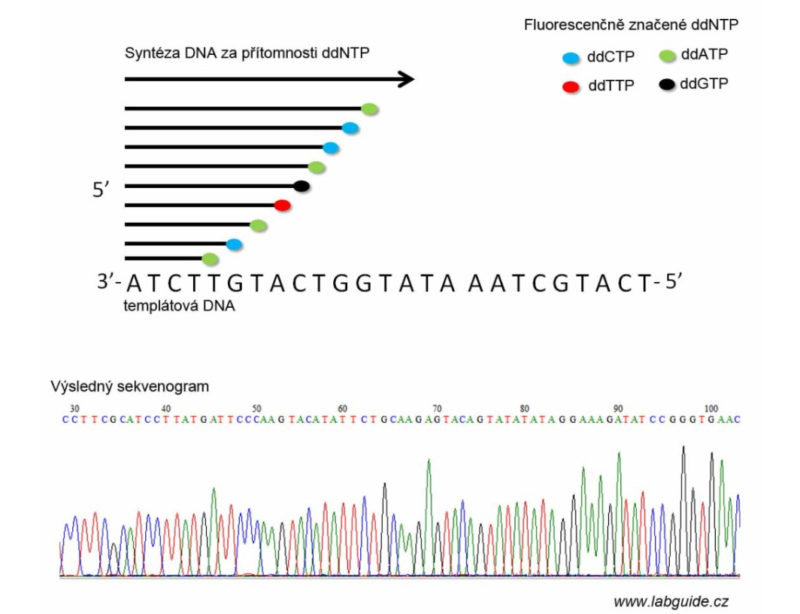
Praktické využití Sangerovy metody
V kapilárovém provedení je Sangerova metoda v současné době stále nejpoužívanější a nejspolehlivější metodou sekvenování. V drtivé většině případů jsou vzorky sekvenovány komerčně specializovanými pracovišti, kdy je pracovišti dodána templátová DNA a případně primery. Při kapilárním sekvenování jsou sekvenovány jednotlivé fragmenty DNA, připravené PCR amplifikací či klonováním, s tím, že při jednom běhu sekvenátoru je obvykle možnost paralelní sekvenace maximálně 96 sekvencí (96 kapilárové sekvenátory). Délka získaných sekvencí je cca 700 bp a tedy maximální sekvenační výtěžek z jednoho běhu sekvenátoru je cca 67 kb. Chybovost Sangerovy metody sekvenování se pohybuje kolem 1,5 % (tzn. jedna báze z cca 66 bází je přečtena chybně).
Literatura:
Maxam AM, Gilbert W (1977). A new method for sequencing DNA. Proc. Natl. Acad. Sci. U.S.A. 74 (2): 560–4.
Sanger F, Coulson AR (1975). A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J. Mol. Biol. 94 (3): 441–8.
Smith LM, Sanders JZ, Kaiser RJ et al. (1986). Fluorescence detection in automated DNA sequence analysis. Nature 321 (6071): 674–9.